

 Дипломная работа: Разработка методов и средств реабилитации объектов отравляющих веществ
Дипломная работа: Разработка методов и средств реабилитации объектов отравляющих веществ
Рисунок 5 - Номенклатура (перечень) гуминовых веществ [30]
Неорганическая часть, органическое вещество и живые организмы принимают участие в процессах самоочищения почвы.
Классификацию почв по степени загрязнения проводят по ПДК химических веществ и их фоновому загрязнению [31]. По степени загрязнения почвы подразделяются на: сильнозагрязненные, среднезагрязненные, слабозагрязненные.
К сильнозагрязненным относят почвы, в которых содержание загрязняющих веществ в несколько раз превышает ПДК, имеющие низкую биологическую продуктивность, существенное изменение физико-химических, химических и биологических характеристик.
К среднезагрязненным относят почвы, в которых установлено превышение ПДК без существенных изменений в свойствах почв.
К слабозагрязненным относят почвы, в которых содержание химических веществ не превышает ПДК, но выше естественного фона.
Коэффициент концентрации загрязнения почвы НС вычисляется по формуле:
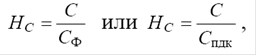
где С – общее содержание загрязняющих веществ;
Сф – среднее фоновое содержание загрязняющих веществ;
Спдк – предельно-допустимое содержание загрязняющих веществ.
Степень устойчивости почвы к химическим ЗВ оценивают по отношению к конкретному токсиканту или группе веществ, которыми загрязнена исследуемая почва. По степени устойчивости к химическим загрязняющим веществам и по характеру ответных реакций почвы подразделяют на: очень устойчивые; среднеустойчивые; малоустойчивые.
Степень устойчивости почвы к химическим загрязняющим веществам характеризуется следующими основными показателями: гумусного состояния почв, кислотно-основными свойствами; окислительно-восстановительными свойствами, катионно-обменными свойствами, биологической активности, уровня подземных вод, доли веществ в почве, находящихся в растворимой форме.
1.4 Микроорганизмы-деструкторы фосфорорганических соединений
Способность микроорганизмов использовать фосфорорганические соединения с С–Р связью в качестве единственного источника фосфора известна сравнительно давно [32]. Впервые доказательство биологического расщепления С–Р связи было получено на примере E. coli, которая в качестве единственного источника фосфора использовала метилфосфоновую или этилфосфоновую кислоты.
Анализ опубликованных к настоящему времени работ свидетельствует о наличии в природе широкого круга микроорганизмов – деструкторов фосфонатов, среди которых грам-положительные и грам-отрицательные бактерии, а также некоторые дрожжи и грибы [33-36]. Однако предполагается, что разлагать фосфонаты способны, скорее, только особые штаммы, но не определенные группы микроорганизмов [37]. Попытка проверить это была предпринята в ряде исследований [33]. Так, из пяти почвенных изолятов были выделены бактерии, способные деградировать широкую группу структурно различных фосфонатов. Прежде всего, это различные штаммы Pseudomonas и Bacillus megaterium, разлагающие 14 из 15 исследованных субстратов, что сопоставимо с широкой субстратной специфичностью, выявленной ранее для Agrobacterium radiobacter. В этой работе впервые было показано, что грам-положительные бактерии могут осуществлять прямое разложение связи С–Р. Способность расти на различных природных и ксенобиотических фосфонатах, как единственных источниках фосфора была проанализирована так же у микроорганизмов из семи экосистем и у 19 лабораторных микроорганизмов. Это исследование показало присутствие деструкторов фосфонатов среди различных бактериальных видов и систематических групп, выделенных как из загрязненных, так и из незагрязненных фосфонатами источников окружающей среды. Это свидетельствует о более широком их распространении, чем предполагалось ранее [34]. Было подтверждено отсутствие способности к деградации фосфонатов эукариотическими организмами. Впервые обнаружена такая способность у фотосинтетического организма Rhodobacter capsulatus. Недавно были выявлены галофильные бактерии Chromohalobacter marismortui, которые способны использовать для роста в качестве единственного источника фосфора фосфоноацетат, 2-аминоэтил-, 3-аминобутил-, метил- и этил- фосфонаты. При этом наблюдались различия в росте клеток на разных фосфонатах и их различная утилизация в зависимости от концентрации NaCl, что, возможно, является результатом использования клетками для этой цели различных транспортных систем. Термофильные бактерии Geobacillus caldoxylosilyticus T20 способны также использовать некоторые фосфонаты в качестве единственного источника фосфора для роста при 600С. Некоторые штаммы Streptomycetes способны использовать в качестве единственного источника фосфора самые различные по структуре фосфонаты. Штамм Streptomyces morookaensis DSM 40565 способен расщеплять 2-амино-4-фосфонобутират в качестве источника азота и фосфора способом, подобным стереоселективному. Эти результаты позволяют предположить наличие нового метаболического пути для расщепления С–Р связи.
Только микроорганизмы, разлагающие алкилфосфонаты с образованием алканов, можно с уверенностью отнести к микроорганизмам, содержащим С–Р лиазу, хотя их субстратная специфичность может и не ограничиваться алкилфосфонатами. Некоторые из указанных микроорганизмов способны разлагать и другие фосфонаты, в том числе глифосат и 2-АЕР (таблица 5).
Большинство бактерий, для которых точно установлено разложение фосфонатов по С–Р лиазному механизму, относится к грам-отрицательным бактериям, однако, известны и два представителя грам-положительных бактерий: Arthrobacter sp. GLP-1 и B. megaterium. Среди почти сорока идентифицированных почвенных изолятов и коллекционных штаммов бактерий, проверенных на С–Р лиазную активность, только вышеупомянутые грам-положительные бактерии обладали ею. У других исследованных грам-положительных бактерий С–Р лиазную активность обнаружить не удалось. Еще одним представителем бактерий, разлагающим фосфонаты по С–Р лиазному механизму, является Rhodobacter capsulatus, фототрофная грам-отрицательная бактерия, способная разрушать С–Р связь при анаэробных условиях на свету. Поиск С–Р лиазной активности среди представителей грибов (Cladosporium herbarum, Fusarium culmorum и Trichoderma viride /33/ не дал положительных результатов.
Таблица 5 - Микроорганизмы, разлагающие фосфонаты по С-Р лиазному механизму
| Микроорганизм | Утилизируемый фосфонат | ||||
| Pn | 2-AEP | Ф-P | глифосат | ||
| Agrobacterium radiobacter | + | + | + | + | |
| Kluyvera ascorbata | + | + | + | - | |
| Kluyvera cryorescens | + | + | + | - | |
| Klebsiella oxytoca | + | + | + | - | |
| Klebsiella pneumoniae | + | + | + | - | |
| Bacillus megaterium 2BLW | + | + | + | + | |
| Arthrobacter sp.GLP-1 | + | + | + | + | |
| Rhizobium sp. | + | + | + | - | |
| Pseudomonas testosteroni DSM 1622 | + | + | - | - | |
| Pseudomonas sp. 7NSK2 | + | + | + | - | |
| Pseudomonas sp. PG2982 | + | + | + | + | |
| Escherichia coli DSM1576 | + | + | - | - | |
| Alcaligenes eutrophus | + | + | + | + | |
| Rhodobacter capsulatus | + | + | + | - | |
| Klebsiella aerogenes | + | + | + | - | |
| Enterobacter aerogenes | + | + | + | слабо | |
Таким образом, способность разлагать фосфонаты по С–Р лиазному механизму встречается среди грам-отрицательных бактерий гораздо чаще, чем среди грам-положительных бактерий. Это отдельные представители классов Arthrobacteriaceae, Bacillaceae, Rhodobacteriaceae, Alcaligenaceae, Pseudomonadaceae, Enterobacteriaceae (Escherichia, Enterobacter, Klebsiella, Kluyvera) и Rhizobiaceae (Rhizobium, Agrobacterium).
Страницы: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10
